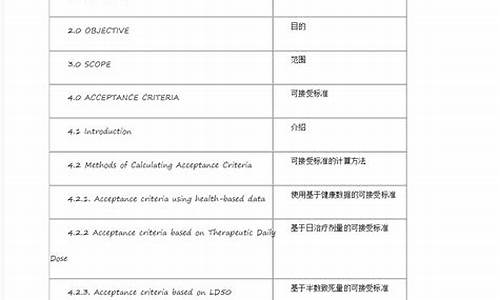
吖啶结构式-吖啶酮结构

DNA聚合酶(DNA dependent DNA polymerase, DDDP): ⑴种类和生理功能:在原核生物中,目前发现的DNA聚合酶有三种,分别命名为DNA聚合酶Ⅰ(pol Ⅰ),DNA聚合酶Ⅱ(pol Ⅱ),DNA聚合酶Ⅲ(pol Ⅲ),这三种酶都属于具有多种酶活性的多功能酶。pol Ⅰ为单一肽链的大分子蛋白质,具有5'→3'聚合酶活性、3'→5'外切酶活性和5'→3'外切酶的活性;其功能主要是去除引物、填补缺口以及修复损伤。pol Ⅱ具有5'→3'聚合酶活性和3'→5'外切酶活性,其功能 不明。pol Ⅲ是由十种亚基组成的不对称二聚体,具有5'→3'聚合酶活性和3'→5'外切酶活性,与DNA复制功能有关。 在真核生物中,目前发现的DNA聚合酶有五种。其中,参与染色体DNA复制的是pol α(延长随从链)和pol δ(延长领头链),参与线粒体DNA复制的是pol γ,polε与DNA损伤修复、校读和填补缺口有关,pol β只在其他聚合酶无活性时才发挥作用。 ⑵DNA复制的保真性:为了保证遗传的稳定,DNA的复制必须具有高保真性。DNA复制时的保真性主要与下列因素有关:①遵守严格的碱基配对规律;②在复制时对碱基的正确选择;③对复制过程中出现的错误及时进行校正。 DNA损伤修复-简史 1949年A.凯尔纳偶然发现灰色链丝菌等微生物经紫外线(UV)照射后如果立即暴露在可见光下则可减少亡。此后在大量的微生物实验中都发现了这种现象,并证明这是许多种微生物固有的DNA损伤修复功能,并把这一修复功能称为光复活。1958年R.L.希尔证明即使不经可见光的照射,大肠杆菌也能修复它的由紫外线所造成的DNA损伤,而后又证明其他微生物也有这种功能,当时就把这种修复功能称为暗复活或暗修复。此后发现暗修复普遍地存在于原核生物、低等真核生物、高等真核生物的两栖类乃至哺乳动物中,并证实暗修复包括切除修复和复制后修复两种。1968年美国学者J.E.克利弗首先发现人类中的常染色体隐性遗传的光化癌变疾病——着色性干皮病(XP)是由基因突变造成的 DNA损伤切除修复功能的缺陷引起的。这一发现为恶性肿瘤的发生机理提供了一个重要的分子生物学证据,也使DNA损伤修复的研究进入了医学领域。 DNA损伤修复-损伤类型 DNA分子的损伤类型有多种。UV照射后DNA分子上的两个相邻的胸腺嘧啶 (T)或胞嘧啶(C)之间可以共价键连结形成环丁酰环,这种环式结构称为二聚体。胸腺嘧啶二聚体的形成是 UV对DNA分子的主要损伤方式。 Χ射线、γ射线照射细胞后,由细胞内的水所产生的自由基既可使DNA分子双链间氢键断裂,也可使它的单链或双链断裂。化学物中的博莱霉素、甲基磺酸甲烷等烷化剂也能造成链的断裂。 丝裂霉素C可造成DNA分子单链间的交联,这种情况常发生在两个单链的对角的鸟嘌呤之间。链的交联也往往带来DNA分子的断裂。 DNA分子还可以发生个别碱基或核苷酸的变化。例如碱基结构类似物5-溴尿嘧啶等可以取代个别碱基,亚硝酸能引起碱基的氧化脱氨反应,原黄素(普鲁黄)等吖啶类染料和甲基氨基偶氮苯等芳香胺致癌物可以造成个别核苷酸对的增加或减少而引起移码突变(见基因突变)。 一种 DNA损伤剂往往可以同时引起几种类型的损伤,其损伤效应的大小和类型与剂量及细胞所处的周期状态有关。 DNA损伤修复-修复方式 光复活又称光逆转。这是在可见光(波长3000~6000埃)照射下由光复活酶识别并作用于二聚体,利用光所提供的能量使环丁酰环打开而完成的修复过程。光复活酶已在细菌、酵母菌、原生动物、藻类、蛙、鸟类、哺乳动物中的有袋类和高等哺乳类及人类的淋巴细胞和皮肤成纤维细胞中发现。这种修复功能虽然普遍存在,但主要是低等生物的一种修复方式,随着生物的进化,它所起的作用也随之削弱。 切除修复 又称切补修复。最初在大肠杆菌中发现,包括一系列复杂的酶促DNA修补复制过程,主要有以下几个阶段:核酸内切酶识别DNA损伤部位,并在5’端作一切口,再在外切酶的作用下从5’端到3’端方向切除损伤;然后在 DNA多聚酶的作用下以损伤处相对应的互补链为模板合成新的 DNA单链片断以填补切除后留下的空隙;最后再在连接酶的作用下将新合成的单链片断与原有的单链以磷酸二酯链相接而完成修复过程。 切除修复并不限于修复嘧啶二聚体,也可以修复化学物等引起的其他类型的损伤。从切除的对象来看,切除修复又可以分为碱基切除修复和核苷酸切除修复两类。碱基切除修复是先由糖基酶识别和去除损伤的碱基,在DNA单链上形成无嘌呤或无嘧啶的空位,这种空缺的碱基位置可以通过两个途径来填补:一是在插入酶的作用下以正确的碱基插入到空缺的位置上;二是在核酸内切酶的催化下在空位的5’端切开DNA链,从而触发上述一系列切除修复过程。对于各种不同类型的碱基损伤都有特异的糖基酶加以识别。不同的核酸内切酶对于不同类型损伤的识别也具有相对的特异性。 切除修复功能广泛存在于原核生物和真核生物中,也是人类的主要修复方式,啮齿动物 (如仓鼠、小鼠)先天缺乏切除修复的功能。 1978年美国学者 J.L.马克斯发现真核生物与原核生物间由于染色质结构不同, 切除修复的过程也不相同。真核生物的DNA分子不象原核生物那样是裸露的,而是缠绕在组蛋白上形成串珠状的核小体结构。真核生物中的嘧啶二聚体的切除分两个阶段:快速切除期,约需2~3小时,主要切除未与组蛋白结合的DNA部分的损伤;缓慢切除期,至少要持续35小时而且需要有某种控制因子去识别这种损伤,使DNA受损部分从核小体中暴露出来,然后经过一系列步骤完成切除修复,然后修复的DNA分子再缠绕在组蛋白上重新形成核小体。 重组修复 重组修复从 DNA分子的半保留复制开始,在嘧啶二聚体相对应的位置上因复制不能正常进行而出现空缺,在大肠杆菌中已经证实这一DNA损伤诱导产生了重组蛋白,在重组蛋白的作用下母链和子链发生重组,重组后原来母链中的缺口可以通过DNA多聚酶的作用,以对侧子链为模板合成单链DNA片断来填补,最后也同样地在连接酶的作用下以磷酸二脂键连接新旧链而完成修复过程。重组修复也是啮齿动物主要的修复方式。重组修复与切除修复的最大区别在于前者不须立即从亲代的DNA分子中去除受损伤的部分,却能保证DNA复制继续进行。原母链中遗留的损伤部分,可以在下一个细胞周期中再以切除修复方式去完成修复。 重组修复的主要步骤有: 1.复制 含有TT或其他结构损伤的DNA仍然可以正常的进行复制,但当复制到损伤部位时,子代DNA链中与损伤部位相对应的位置出现切口,新合成的子链比未损伤的DNA链要短。 2.重组 完整的母链与有缺口的子链重组,缺口由母链来的核苷酸片段弥补。 3.再合成 重组后母链中的缺口通过DNA多聚酶的作用合成核酸片段,然后由连接酶是新片段与旧链连接,至此重组修复完成。 重组修复并没有从亲代DNA中去除二聚体。当第二次复制时,留在母链中的二聚体仍使复制不能正常进行,复制经过损伤部位时所产生的切口,仍旧要用同样的重组过程来弥补,随着DNA复制的继续,若干代以后,虽然二聚体始终没有除去,但损伤的DNA链逐渐“稀释”,最后无损于正常生理功能,损伤也就得到了修复。 SOS修复 是SOS反应的一种功能。SOS反应是DNA受到损伤或脱氧核糖核酸的复制受阻时的一种诱导反应。在大肠杆菌中,这种反应由recA-lexA系统调控。正常情况下处于不活动状态。当有诱导信号如 DNA损伤或复制受阻形成暴露的单链时,recA蛋白的蛋白酶活力就会被激活,分解阻遏物lexA蛋白,使SOS反应有关的基因去阻遏而先后开放,产生一系列细胞效应。引起SOS反应的信号消除后,recA蛋白的蛋白酶活力丧失,lexA蛋白又重新发挥阻遏作用。 SOS 反应发生时, 可造成损伤修复功能的增强。如uvrA、uvrB、uvrC、uvrD、ssb、recA、recN和ruv基因发达从而增强切除修复、复制后修复和链断裂修复。而recA和umuD.C则参与一种机制不清的易错修复,使细胞存活率增加,突变率也增加。 除修复作用外,SOS反应还可造成细胞分裂受阻、溶原性噬菌体释放和DNA复制形式的改变。后者指DNA聚合酶I*的形成,使DNA复制的准确性降低并可通过损伤部位。此时,DNA复制的起始也无需新合成蛋白。 在真核细胞中,虽然还不清楚具体过程,但肯定存在可诱导的易错修复。酵母RAD6系统就是一种易错修复系统。在哺乳类细胞中,DNA损伤可诱导细胞内病毒的释放、病毒转化作用的加强、染色体重组增强和细胞纤溶酶激活物的形成等。并且还发现了和大肠杆菌相似的ω-复活效应和ω-诱变效应。由于这种反应可增强突变、染色体重排和病毒的活动,以及对 DNA复制形式的影响,可能与癌基因激活和肿瘤形成有直接的关系。因而,SOS反应可作为检测药物致癌性的指标,而抑制SOS反应的药物则可减少突变和癌变。这类物质被称之为抗变剂。 适应性修复 1977年美国学者L.萨姆森等在大肠杆菌中发现的不同于 SOS修复的又一种诱导反应,它可以修复鸟嘌呤碱基的甲基化。如先以每毫升培养基 1微克的诱变剂N-甲基-N'-硝基-亚硝基胍 (MNNG)培养大肠杆菌两小时,就能使大肠杆菌对MNNG浓度高几百倍的环境产生抗性。这是由于 MNNG引起的DNA链上的鸟嘌呤甲基化诱导合成甲基受体蛋白,这种甲基受体蛋白分子的半胱氨酸能和甲基基团结合形成S-甲基半胱氨酸,从而使甲基化的鸟嘌呤碱基得以修复。 链断裂修复 包括DNA分子的单链断裂修复、双链断裂修复和染色体的断裂重接修复。在连接酶的参与下这些断裂能够迅速地以重接的方式修复。这种修复有两个特点:一是不稳定性,重接后又可以再度离解;二是不正确性,经常发生随机的重接错误。 链交联修复 起始步骤是在糖基酶的催化下解开交联的一条臂, 通过碱基切除的方式先修复合成其中一条单链,然后再在内切酶的催化下,以核苷酸切除修复的方式从相反的方向修复对侧的单链片断。 DNA损伤修复-检测方法 大部分DNA损伤修复都依赖于DNA的修复合成,所以对修复合成的测定常用来作为DNA修复的检测方法。常用的有以下几种: 放射自显影法 在细胞培养物中加入氚标记的胸腺嘧啶核苷等放射源,用放射自显影方法计数银颗粒数来测定修复合成过程中参入到DNA分子中的量。 液体闪烁计数法 用液体闪烁计数器测定培养物中的放射源因修复合成而参入到DNA分子中的量。这一方法适用于大批量样本。 超速离心法 一种应用比较广泛的方法,可应用于切除修复、复制后修复及链断裂修复方式的检测。一般是用氚标记溴脱氧尿嘧啶核苷等参入到修复合成的DNA分子中去以改变DNA分子的重量(BrdU的分子量比尿嘧啶核苷大),通过超速离心可以从沉降系数不同的各组分中收集修复合成中参入量不同的DNA片断,然后分别测定其放射性的强度来判断修复合成的多少。 病毒宿主细胞复活法 以SV40病毒、腺病毒、疱疹病毒、噬菌体等感染培养的人体细胞或细菌,然后以紫外线等处理以造成病毒DNA分子的损伤,因为病毒DNA分子损伤的修复是靠宿主细胞的修复酶系统,所以受损伤的病毒能否继续生存繁殖可间接地反映宿主细胞的修复功能。 姐妹染色单体互换(SCE)法 姐妹染色单体互换率的检测也能反映一部分DNA修复功能。人类中的某些先天性DNA修复缺陷疾病如布卢姆氏综合征患者的自发SCE显著增高;另一些如着色性干皮病则诱发SCE增高。这是由于DNA修复功能的缺陷导致染色体稳定性减弱所致。 实践意义DNA修复与肿瘤各种原因引起的DNA损伤可以通过各种方式修复。如果修复功能有缺陷,DNA损伤就可能造成两种结果:一是细胞亡;二是发生基因突变,或进而恶性转化为肿瘤细胞。先天性DNA修复缺陷疾病患者容易发生各种恶性肿瘤,例如人类的着色性干皮病患者的皮肤对阳光过度敏感, 照射后出现红斑、水肿,继而出现色素沉着、干燥、角化过度,结果可导致黑色素瘤、基底细胞癌、鳞状上皮癌及棘状上皮瘤的发生。通过细胞融合的研究表明具有不同临床表现的该病患者有明显的遗传异质性,可以分为A、B、C、D、E、F、G七个互补群及变种,A-G互补群表现为不同程度的核酸内切酶缺乏引起的切除修复功能缺陷,变种的切除修复功能正常,但复制后修复的功能有缺陷。又如范可尼贫血临床主要表现的特征如再生障碍性贫血、生长迟缓、易患白血病等是由于先天性链交联等修复缺陷所致。其他如布卢姆氏综合征和毛细血管扩张共济失调患者都易患白血病和淋巴肉瘤,也是先天性DNA修复缺陷造成的。 值得注意的是DNA修复功能缺陷虽可引起肿瘤的发生,但已癌化细胞本身的DNA修复功能并不低下,相反地却显著地升高,并能够充分地修复化疗药物引起的DNA损伤, 这也是大多数抗癌药物不能奏效的原因。地鼠细胞的DNA损伤修复的方式以复制后修复为主, 如果在地鼠的浆细胞瘤细胞的培养物中加入环磷酰胺等抗癌药后,瘤细胞照样生长,如果加入环磷酰胺的同时再加入咖啡因(复制后修复的抑制剂),则瘤细胞的生长受到了明显的抑制。所以DNA修复的研究可为肿瘤联合化疗提供方案。 DNA损伤修复-DNA修复与衰老 从DNA修复功能的比较研究中发现寿命长的动物(象、牛等)修复功能较强;寿命短的动物 (仓鼠、小鼠、鼩鼱等)修复功能较弱。人的DNA修复功能也很强,但到一定年龄后逐渐减弱,同时突变细胞数也相应增加,所以老年人癌的发病率也比较高。检测各年龄组正常人的染色体畸变率和 DNA修复功能证实了这一点。人类中常染色体隐性遗传的早老症和韦尔纳氏综合征患者一般早年于心血管疾病或恶性肿瘤;患者的体细胞极易衰老,是研究老年病与DNA修复关系的很好模型。 DNA修复与免疫 DNA修复功能先天缺陷的病人的免疫系统也常是有缺陷的,主要是 T淋巴细胞功能的缺陷。随着年龄的增长细胞中的DNA修复功能逐渐衰退,如果同时发生免疫监视机能的障碍,便不能及时清除癌化的突变细胞,从而导致发生肿瘤。所以, 衰老、DNA修复、免疫和肿瘤四者是紧密关联的。 DNA损伤修复-DNA修复与环境致癌因子的检测 DNA修复的研究已被应用于检测各种化学致癌物。一般的方法是在体外传代培养的正常人皮肤成纤维细胞或大鼠原代培养的肝细胞中加入被检物,培养一定时间后再加入继续培养,然后收集细胞作放射自显影或液体闪烁的测试,如果参入量显著增高,表明被检物可疑为诱变剂或致癌剂。微生物培养的方法则更为简便、迅速,例如可以用枯草杆菌重组功能发生缺陷的突变型来进行检测,这些突变型由于丧失了重组功能而不能进行重组修复,因而更容易为许多诱变剂和致癌剂所杀伤致。 关于DNA修复机制方面的许多问题还有待于进一步的研究阐明。例如从原核生物开始到真核生物的高等哺乳类动物各依靠哪些方式来修复受损伤的DNA分子,修复方式又是怎样随物种的进化而发生演变的,修复缺陷的遗传异质性的本质又是什么,免疫缺陷和DNA修复功能缺陷的因果关系又是怎样的等等。
5碳环叫茂,6碳环叫苯
苯胺的结构是:苯(上下结构)胺(左右结构)。
苯胺的结构是:苯(上下结构)胺(左右结构)。注音是:ㄅㄣˇㄢ_。词性是:名词。拼音是:běnàn。
苯胺的具体解释是什么呢,我们通过以下几个方面为您介绍:
一、词语解释点此查看计划详细内容
苯胺běn’àn。(1)一种油状有毒液体胺C6H5NH2,纯品无色,可由(例如靛蓝或煤的)毁馏制得,但是现在常用还原硝基苯或氯苯和氨高压反应制得,主要用于有机合成(例如染料、药物、橡胶、化学试剂和炸药)和作溶剂;氨基苯。
二、网络解释
苯胺苯胺又称阿尼林、阿尼林油、氨基苯,分子式:C6H7N。无色油状液体。熔点-6.3℃,沸点184℃,相对密度1.02(20/4℃),相对分子量93.128,加热至370℃分解。稍溶于水,易溶于乙醇、等有机溶剂。苯胺是最重要的胺类物质之一。主要用于制造染料、药物、树脂,还可以用作橡胶硫化促进剂等。它本身也可作为黑色染料使用。其衍生物甲基橙可作为酸碱滴定用的指示剂。2017年10月27日,世界卫生组织国际癌症研究机构公布的致癌物清单初步整理参考,苯胺在3类致癌物清单中。
关于苯胺的成语
东拉西扯促膝谈心斗粟尺布黩武穷兵电光朝露担惊受怕独步当时岛瘦郊寒独辟蹊径惮赫千里
关于苯胺的词语
大含细入黩武穷兵点金乏术电光朝露惮赫千里独辟蹊径东拉西扯词不达意担惊受怕斗粟尺布
关于苯胺的造句
1、喹啉加氢生成四氢喹啉,四氢喹啉进一步裂化转化为吡啶、苯胺和氨。
2、所述基质可以包括胶原蛋白、脱乙酰壳多糖或氧化再生纤维素,并且所述染料例如为苯胺或吖啶染料。
3、在两相成均相后,将混合物在控制速率下冷却,则两相再分离的温度即为苯胺点或样品的混合苯胺点。
4、以苯胺、邻硝基氯苯为原料,采用直接缩合法引入相转移催化剂合成邻硝基二苯胺。
5、苯胺也叫氨基苯、阿尼林油,是制造染料、橡胶助剂、塑料、农药、医药的主要原料,可作炸药中的稳定剂、汽油中的防爆剂以及溶剂。
点此查看更多关于苯胺的详细信息
细胞分裂早中期、前中期、晚前期或更早时期染色体的带纹,称为什么?
楼上瞎扯,架乙丙这些,只适合链烃和脂肪环烃。而杂环化合物因为存在大派键命名方式并不相同。
最常见的杂环化合物是五元和六元杂环及苯并杂环化合物等。五元杂环化合物有:呋喃、噻吩、吡咯、噻唑、咪唑、唑等。六元杂环化合物有:吡啶、吡嗪、嘧啶等。稠环杂环化合物有:吲哚、喹啉、蝶啶、吖啶等。杂环化合物中,最小的杂环为三元环,最常见的是五、六元环,其次是七元环。
杂环化合物常以俗名命名,较少用系统命名。系统命名是指以相应的碳环为母体而命名。例如,含两个不饱和键的环戊二烯称为茂,与之相应的一种杂环化合物,例如吡咯,可以看成是由NH取代了茂中的CH2而成 ,称为氮(杂)茂。依此类推,吡啶称为氮(杂)苯,喹啉称为氮(杂)萘等,但一般仍习惯于用俗名命名。
杂环化合物的中文名称是以口字旁标明其为杂环,另半部分表明杂原子的种类。例如,以?、噻分别表示为含氧、硫的杂环;以咯、唑、嗪、啶、啉表示为含氮的杂环,这些字是根据英文字的尾音创造的,其中咯、唑表示为五元含氮杂环,其余的指六元含氮杂环。杂原子超过一个者分别以二、三等字表示相同杂原子的数目,例如?二唑,表示该杂环化合物为含有一个氧和两个氮杂原子的五元杂环。在环中不同的原子可有不同的排列方式,命名时各原子的位置编号遵循下列原则:①只含一个杂原子或一个以上相同杂原子的杂环,杂原子编最小号。②含两个不同杂原子时,不同杂原子的编号顺序为氧、硫、氮,例如 1,2,3-?二唑,氧在第一位,两个氮原子分别在第二和第三位 ,其余为碳原子的五元杂环化合物。
卢沟桥为什么“成了我国人民永远难忘的一处具有历史意义的建筑”?
带型(带型)
该染色体带型。随着细胞学特殊的处理呈现出不同色调的染色,染色体。染色的数量,位置,宽度和颜色深度,是相对稳定的,所以每个染色体带型模式,被称为固定。染色体带型的染色体鉴定的重要依据。通过区划机制,染色体组成,结构,行为和功能的许多信息都可以得到。的染色体的研究工作始于20世纪60年代中后期。染色体显带技术是治疗的经理因素,染色体染色呈现出不同的色调与一个特定的模式的方法,也被称为显带技术。一般细胞学染色,染色体的着色是均匀的。治疗子带技术,反映了染色体的固有结构,在染色体上的频带,并可以显示不同的染色体上的属同一物种的差异的的物种差异或不同的染色体。常见的条纹与显示技术的Q-带,G带,C带,R带,T区。对于每个子带的技术,每个染色体的带型的是一个非常具体的和恒定的。 Q-波段技术是1968年在瑞典的细胞的的化学家Kasiposong(T.Caspersson),显示芥子喹吖啶酮染色体染色在紫外光照射下的荧光带,这些区域的DNA分子,在AT基地丰富的部分的成分。 ,姬姆萨带G的,将在该区域的中期细胞胰蛋白酶或碱,热,尿素处理后,再由姬姆萨染料染色的介绍后,。 C波段,也被称为着丝粒异染色质(MLPardue),在1970年建立,中期染色体后,通过盐酸碱(如氢氧化钡)处理,然后Giemsa染色,显示相邻的着丝粒异染色质。 R区是染色体盐酸水解或没有的情况下,胰蛋白酶处理区提出后,姬姆萨染色,显示G带染色带颜色的区域,所谓的反带。 T带,也被称为端粒带区是由染色体的端粒吉姆萨和吖啶橙染色,典型的T与绿色份。 20世纪70年代末,由于细胞的同步和显带技术的改进的方法,从而获得更长的时间,而与图案更丰富的染色体,这被称为一个高分辨率的染色体染色体。例如,1975年以后,美国细胞遗传学家龙尼斯(JJRon待业待学青少年)建立一个高分辨率的显带方法与氨甲喋呤,第一次细胞分裂同步,然后秋季酰胺很短的时间,使其出现了大量的晚预和早中期师的。早期的染色体显带染色体阳性的长度比,可以做出来的分区细染色体带型。例如,前中期显示555至842与843至1256与晚前3000至10,000区分程度较高的带式所示,从早早期较长的染色体获得的。染色体,其畸变的高分辨率技术可以提供更多的细节,以帮助找出更细微的染色体异常染色体断点,定位更准确,这些详细的基因图谱的显著值的映射。总之,无论是在医疗诊断中的细胞遗传学和遗传学,动物和植物育种,带技术的理论研究中广泛使用的技术。
喹啉结构式
因为中国抗日军队在卢沟桥打响了全面抗战的第一枪。
金大定二十九年六月,卢沟桥始建;1961年卢沟桥被公布为第一批国家重点文物保护单位;1985年卢沟桥正式退役;1991年卢沟桥实现封闭管理。
卢沟桥全长266.5米,宽7.5米,桥两侧雁翅桥面呈喇叭口状。1937年7月7日日本在此发动全面侵华战争,史称“卢沟桥事变”(亦称“七七事变”)。 中国抗日军队在卢沟桥打响了全面抗战的第一枪。
卢沟桥的建设成果:
卢沟桥的半圆拱券采用纵联式实腹砌筑法,使11个拱券联成一体。拱券龙门石上,2019年依然保留有中间三个被称作虬蠖的拱顶龙头,雕工十分精美。
拱券石块之间都用铁铜子或铁件联结加固,桥墩内部也都用铁活上下拉联,桥脚以铁柱穿石,俾使千载永固。这在中国大石桥建筑史上也是罕见的。
细菌体内的遗传物质有哪两种
喹啉的结构式为C9H7N,是一种由一个苯环和一个氮原子组成的六元杂环化合物。
喹啉,是一种重要的有机化合物,属于氮杂环己烷类。其结构式由一个苯环和一个氮原子组成,形成一个六元杂环。喹啉的分子式为C9H7N,每个碳原子均以sp2杂化轨道与其它原子形成化学键。
在喹啉分子中,氮原子替代了苯环中的一个碳原子。这个氮原子具有一对孤对电子,使得喹啉具有芳香性。这使得喹啉在化学反应中具有独特的性质,例如它能与多种金属离子结合生成络合物,并在许多有机反应中作为配体。
喹啉环具有多个活性位点,可以进行多种化学反应,如亲电取代、亲核取代、氧化、还原等。这些反应可用来合成各种复杂的化合物,包括药物、农药、染料等。
喹啉的结构式是一个重要的有机化学概念,它代表了一种具有特殊结构和反应性能的化合物。这种化合物的存在和性质对于有机化学、药物化学和材料科学等领域的发展具有重要意义。通过深入了解喹啉的结构式和反应机理,我们可以更好地利用这种化合物来创造更多有价值的物质。
喹啉的用途:
1、药物合成:喹啉及其衍生物具有多种生物活性,可用于合成多种药物,如抗菌药、抗炎药、抗肿瘤药等。由于喹啉环具有芳香性,它可以与许多药物分子结合,从而提高药物的疗效和稳定性。
2、农药合成:喹啉也是一种重要的农药合成原料,可以用于合成多种杀虫剂、除草剂、杀菌剂等。通过在喹啉环上引入不同的取代基或连接不同的侧链,可以制备出具有不同作用机制和杀虫谱的农药。
3、染料合成:喹啉环具有较好的共轭体系,因此它可以用于合成各种染料和颜料。例如,喹吖啶酮染料是一种重要的红色染料,可用于纺织品、皮革、纸张等材料的染色。
4、化学分析:喹啉具有荧光性质,可在紫外线下发出蓝绿色的荧光,它可用于荧光分析。此外,喹啉还可以作为金属离子的配体,用于络合滴定和光度分析等分析方法。
5、工业用途:喹啉在工业上也有广泛的应用,如用作溶剂、化学中间体、催化剂等。由于喹啉具有良好的溶解性和反应性,它可以用于许多化学反应中作为溶剂或反应物。
屠呦呦简历 三分钟演讲
一、细菌染色体
细菌作为原核型微生物,虽没有完整的核结构,但却有核区(或核质)。在电镜下观察,核区有盘旋堆积的DNA纤维。自大肠杆菌提取的DNA是一条完整的DNA链,分子量为 2.4×109daltons,仅为人体胞DNA量的0.1%。细胞的DNA含量决定存在的基因数。如按每个基因由平均为1000个碱基对估计,大肠杆菌的DNA约为4×106个碱基对,因此约有4000个基因,可编码几千种多肽。细菌染色体DNA与其他生物相同,由互补的双链核苷酸组成。细菌的染色体与生物细胞染色体不同,前者不含有组蛋白,基因是连续的,无内含子。由于细菌核区DNA的功能与真核细胞染色体的功能相同,因此又称其为细菌染色体。
二、质粒
细菌的DNA除大部分集中于核质(染色体)内,尚有少部分(约1~2%)存在于染色体外,称为质粒。质粒与染色体的相似处为:质粒亦为双链环形DNA,不过其分子量远比染色体为小,仅为细菌染色体DNA的0.5~3%。质粒亦可携带遗传信息,可决定细菌的一些生物学特性。然而质粒却有一些与染色体DNA 不同的特性。
1.质粒并非细菌生存所必不可少的遗传物质。细菌如失去染色体,则不能生存;然而细菌失去质粒后仍能生存。这是由于染色体DNA携带的基因所编码的产物,在细菌新陈代谢中是生存所必须者;而质粒携带的基因所编码的产物并非细菌的生存所必须者。因此质粒可以在细菌间传递与丢失。
2.质粒的传递(转移)是细菌遗传物质转移的一个重要方式。有些质粒本身即具有转移装置,如耐药性质粒(R质粒);而有些质粒本身无转移装置,需要通过媒介(如噬菌体)转移或随有转移装置的质粒一起转移。获得质粒的细菌可随之而获得一些生物学特性,如耐药性或产生细菌素的能力等。
3.质粒可自行失去或经人工处理而消失。在细菌培养传代过程中,有些质粒可自行从宿主细菌中失去。这种丢失不像染色体突变发生率很低,而是较易发生。用紫外线、吖啶类染料及其他可以作用于DNA的物理、化学因子处理后,可以使一部分质粒消失,称为消除。目前学者们感兴趣的是如何通过人工处理消除耐药质粒或与致病性有关的质粒。
4.质粒可以独立复制。质粒为DNA,有复制的能力,质粒的复制可不依赖于染色体,而在细菌胞浆内进行。这一特性在基因工程中需扩增质粒时很有用处,因可使细菌停止繁殖而质粒仍可继续复制,从而可获得大量的质粒。
5.可有几种质粒同时共存在于一个细菌内。因质粒可独立复制,又能转移入细菌和自然失去,因此就有机会出现几种质粒的共存。但是并非任何质粒均可共存,因发现在有些情况下,两种以上的质粒能稳定地共存于一个菌体内,而有些质粒则不能共存。
目前已在很多种细菌中发现质粒。比较重要者有决定性菌毛的F因子,决定耐药性的R因子以及决定产大肠杆菌素的Col因子等。耐药性质粒的分子量相对较小,而与致性有关的质粒则为大质粒。革兰氏阴性菌一般都带有质粒。某些革兰氏阳性菌如葡萄球菌也有质粒。
屠呦呦,女,药学家。1930年12月30日生于浙江宁波,1951年考入北京大学,在医学院药学系生药专业学习。 1955年,毕业于北京医学院(今北京大学医学部)。毕业后曾接受中医培训两年半,并一直在中国中医研究院(2005年更名为中国中医科学院)工作,期间前后晋升为硕士生导师、博士生导师,现为中国中医科学院的首席科学家。 。中国中医研究院终身研究员兼首席研究员,青蒿素研究开发中心主任,博士生导师、药学家,诺贝尔医学奖获得者。
屠呦呦多年从事中药和中西药结合研究,突出贡献是创制新型抗疟药青蒿素和双氢青蒿素。1972年成功提取到了一种分子式为C15H22O5的无色结晶体,命名为青蒿素。2011年9月,因为发现青蒿素——一种用于治疗疟疾的药物,挽救了全球特别是发展中国家的数百万人的生命获得拉斯克奖和葛兰素史克中国研发中心“生命科学杰出成就奖”。2015年10月,屠呦呦获得诺贝尔生理学或医学奖,理由是她发现了青蒿素,这种药品可以有效降低疟疾患者的亡率。她成为首获科学类诺贝尔奖的中国人。
屠呦呦是第一位获得诺贝尔科学奖项的中国本土科学家、第一位获得诺贝尔生理医学奖的华人科学家。[是中国医学界迄今为止获得的最高奖项,也是中医药成果获得的最高奖项。
研发青蒿素
抗疟新药青蒿素的第一发明人
疟疾是危害严重的世界性流行病,全球百余国家年约三亿多人感染疟疾。自本世纪六十年代起,氯喹等原有抗疟药因疟原虫对此产生抗药性而失效。时值越南战争,促使国际上迫切寻找新结构类型抗疟药。在国内(曾由“523”办组织全国七大省市筛选中草药3200多种,亦有人做过青蒿,认为无效而放弃),国外(美国结合侵越战争需要,筛选化合物达三十万种)大量工作未获成功的情况下,领导科研组以现代科学继承发扬祖国医药学遗产为指导思想,从系统整理历代医籍、本草入手,收集二千多种方药基础上,归纳编篡成《抗疟方药集》,又从中选200多方药,组织筛选。经研制380多个样品,特别结合古代用药经验,从东晋?葛洪《肘后备急方》青蒿“绞汁”服用截疟记载中,考虑到温度、酶解等因素,不断改进提取方法,终于在1971年获得青蒿抗疟发掘成功。
发现的有效部位和有效单体—青蒿素,对鼠疟、猴疟均具有100%的抗疟作用。经大量提取药物,安全性实验及自身试服后于1972年,1973年分别就有效部位和青蒿素单体,在海南昌江疟区作首次临床研究,证实具有胜于氯喹的满意疗效,抗疟新药—青蒿素由此诞生。
屠呦呦等自1972年分离得到青蒿素单体后即着手进行青蒿素的化学结构研究,先后与中国科学院有机所和生物物理所协作,最终用X—衍射方法确定了青蒿素的立体结构。青蒿素是一个具过氧基团的新型倍半萜内酯,也是一个与过去抗疟药结构完全不同的新抗疟药,打破了过去认为“抗疟药必须含氮杂环”的断言。为此青蒿素的发现不仅找到了一个能解决抗性疟疾的新药,而且为寻找新的抗疟药开辟了一条新的途径。经全国协作,验证病例达二千多,确证为“高效、速效、低毒”的抗疟新药,特别对抗氯喹恶性疟有特效。
发现双氢青蒿素
首先发现双氢青蒿素
在研究青蒿素化学结构中,屠呦呦于1973年首创其还原衍生物—双氢青蒿素。这在青蒿素类药物研究上意义是很大的:
(一)确证青蒿素结构中羰基的存在;(二)由双氢青蒿素结构中的羟基得以制备各类青蒿素衍生物,增效并扩大生物活性,促使当今国内、外对青蒿素类药物研究不断深入,更好为人类保健作贡献;(三)经屠呦呦负责的科研组七年努力,得以创制出临床药效高于青蒿素10倍,又复燃率低至1.95%的新一代抗疟药,其片剂具有更突出的“高效、速效、安全、剂量小、口服方便、复燃率低、研制简廉”等优点,特别是经药效学及临床研究,证实最便于病人口服的片剂,在“安全、有效”上,胜于当前注射给药的同类衍生物,被认为是当前青蒿素类药物之优选者!该药于1992年获国家“一类新药证书”(92卫药证字X-66、67),已大批生产,经销国际市场,反映良好,被认为是“国际上治疗各型疟疾的较理想口服治疗药”。
研究青蒿
完成“青蒿品种整理和质量研究”
屠呦呦在2011年度“拉斯克奖”颁奖典礼上
“七五”期间,参与国家攻关项目《常用中药材品种质量研究》中“青蒿品种整理和质量研究”课题,对青蒿进行系统研究,分离鉴定了17个化合物,其中5个为新化合物,并修正了《中国药典》长期沿用的谬误。
为防止青蒿素抗药性的产生,保护青蒿素的临床使用寿命,避免滥用,与首都医科大学合作,就“双氢青蒿素对约氏疟原虫在蚊体内发育的影响”进行研究,发现青蒿素影响约氏疟原虫配子体感染性,但对蚊体内子孢子增值期不起抑制作用,即未能抑制卵囊继续发育到子孢子。提示青蒿素类药物不能用于疟疾预防。
1978年青蒿素鉴定会后,屠呦呦负责的科研组工作继续不断深入,青蒿素获1985年卫生部实施新药审批办法后的第一个《新药证书》(86 -x-01号)。1981年应WHO的请求,在北京召开“青蒿素”专题的国际会议,屠呦呦以“青蒿素的化学研究”为题,第一个作报告,获得高度评价,认为“青蒿素的发现不仅增加一个抗疟新药,更重要的意义还在于发现这一新化合物的独特化学结构,它将为合成设计新药指出方向”。由此带动国际抗疟领域工作的新进展,也促使世界上很多国家对青蒿素展开进一步的研究。
复方双氢青蒿
研制青蒿素类和吖啶类抗疟药组成的“复方双氢青蒿素”
为解决多重抗药性恶性疟蔓延全球的问题,近年国外十分重视复方的研究。双氢青蒿素为当前青蒿素类药物中的首选药,对多重抗药性恶性疟无交叉抗药性。而吖啶类抗疟药—咯萘啶与4-氨基喹啉类、嘧啶类、胍类及磺胺类均无交叉抗药性。为此设计了双氢青蒿素与咯萘啶组成的复方。在军科院支持下,进行实验研究,确证二药具显著协同增效作用,增效指数为7.6,(高于当前广泛应用的蒿甲醚和本芴醇组成的复方蒿甲醚,其增效指数为6),且作用位点多,疗程短。经二药联用治疗恶性疟10例,在12-24小时内,分二次服用,即达高效,40天未见原虫复燃。已获专利证书(专利号:ZL 99109669.X)。
双氢青蒿素栓剂
据WHO资料,每天约有3000个婴幼儿童于疟疾。为此研发了“双氢青蒿素栓剂”,方便儿童直肠给药。又用直接得自青蒿的青蒿素制成口服片剂,制作简便,价格便宜,又不易产生抗药性。均于2003年获得《新药证书》,分别为国药证字H20030341和H20030144。
扩展药效
经研究发现,双氢青蒿素在免疫领域具良好的双向调节作用,既能降低B细胞高反应性以减少免疫复合物沉积所致的自身免疫病,又可提高T细胞的免疫功能。在北医有关部门支持下,我们已将双氢青蒿素用于治疗红斑狼疮和光敏性疾病。现已获国家食品药品监督管理局的“药物临床研究批件”(2004L02089)和中国发明专利(专利号:ZL 99103346.9)。经临床100例疗效初步观察,总有效率94%,显效率44%。
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。